- Teste de hematologie
- Teste de biochimie
- Biochimie generală din sânge și urina
- Proteine specifice in ser si urina
- Teste biochimice din lichide de punctie
- Teste biochimice din materii fecale
- Teste biochimice pentru tulburari ereditare de metabolism
- Teste pentru nefrolitiaza
- Vitamine, oligoelemente, stres oxidativ
- Acizi grași
- Transferina carbohidrat deficitara (CDT) marker pentru alcoolism
- Markeri non-invazivi pentru afecţiunile hepatice
- Analiza chimică calculi
- Markeri endocrini
- Markeri tumorali
- Markeri virali
- Markeri cardiaci
- Markeri anemie
- Markeri ososi
- Markeri boli autoimune
- Anticorpi antispermatozoizi
- Autoanticorpi in afectiuni endocrine, cardiace, renale
- Autoanticorpi in afectiuni neurologice
- Autoanticorpi in afectiunile dermatologice
- Autoanticorpi in anemia pernicioasa
- Autoanticorpi in diabetul zaharat
- Markeri pentru afectiuni hepatice si gastrointestinale autoimune
- Markeri pentru afectiuni reumatismale si vasculite
- Markeri pentru monitorizarea evolutiei si tratamentului
- Markeri pentru sindromul antifosfolipidic
- Serologie boli infectioase
- Teste specializate de alergologie si imunologie
- Teste de biologie moleculara
- Teste de citogenetica
- Teste de microbiologie
- Toxicologie
- Citologie cervico-vaginala
- Histopatologie
- Consult genetic
- Genetica medicala

Gena de fuziune BCR-ABL1-detectie cantitativa
Preț: 1452.00 lei
Analiza medicala Gena de fuziune BCR-ABL1-detectie cantitativa se efectuează conform următoarelor specificații/ condiții:
-
- Centrele de recoltare și laboratoarele din București și zonele limitrofe: recoltarea se efectuează de luni până joi, în timpul programului de recoltare;
- Centrele de recoltare și laboratoarele din țară: recoltarea se efectuează de luni până miercuri, în timpul programului de recoltare.
Informatii generale
Leucemia mieloida cronica (LMC) – prototipul afectiunilor leucemice – constituie o boala clonala a celulei stem din maduva osoasa ce conduce predominant la hiperproliferarea elementelor granulocitare in toate stadiile de maturatie.
LMC se caracterizeaza prin prezenta translocatiei cromozomiale t(9;22)(q34;q11) care a fost considerata initial ca o anomalie 22 q- (un cromozom 22 cu bratul lung scurtat) si denumita cromozomul Philadelphia (Ph) (Nowell, 1960; Rowley, 1973). Aceasta translocatie are drept rezultat juxtapunerea genei tirozin kinazei ABL1 (Abelson) de pe cromozomul 9 langa gena BCR (Breakpoint Cluster Region) de pe cromozomul 22 cu formarea genei de fuziune BCR-ABL1 (fuziune cap-coada capat 5’ BCR – capat 3’ ABL; vezi figura 1). Gena anormala se transcrie intr-un ARNm hibrid ce va determina in final sinteza unei proteine himerice Bcr-Abl cu activitate tirozin-kinazica autonoma3;8.
Figura 1: Cromozomii 9 si 22 normali – translocatia (9,22)
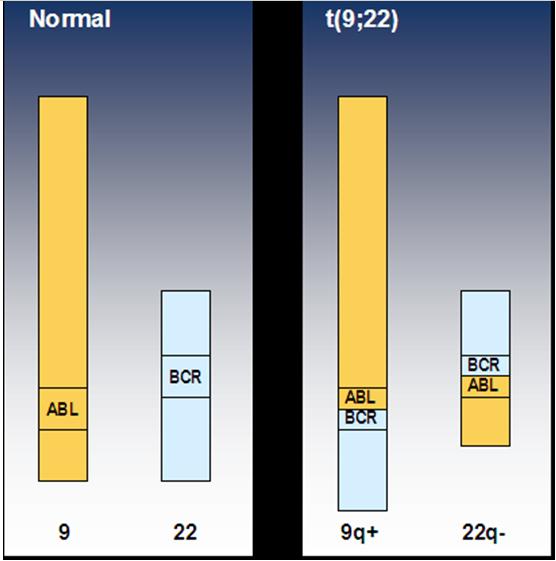
O translocatie identica citogenetic cu cea din LMC poate fi intalnita la aproximativ 20-25% dintre pacientii adulti si la ~5% dintre copiii cu leucemie acuta limfoblastica (LAL), precum si in unele cazuri rare de leucemie acuta mieloblastica (LAM)4;8.
Studiile referitoare la modul in care genele normale BCR si ABL regleaza cresterea au indicat cateva posibilitati prin care molecula himerica ar putea facilita proliferarea celulara necontrolata8.
La persoanele normale proteinele codificate de genele BCR si ABL se exprima virtual in toate celulele3. Gena ABL1 (V-abl Abelson murine leukemia viral oncogene homolog 1) este o protooncogena ce codifica o tirozin kinaza Abl1 cu o greutatea moleculara de 145 kD care isi fosforileaza reziduurile de tirozina proprii (autofosforilare) si pe cele ale altor proteine. Produsul de transcriptie are fie 6, fie 7 kb, in functie de modul de splicing alternativ.
Atunci cand capatul N- terminal al proteinei este codificat de exonul 1 (ABL1A), proteina este localizata in nucleu; cand este implicat exonul 1b (ABL1B), conformatia glicinei N –terminale este modificata de catre acidul miristic, ceea ce duce la directionarea proteinei spre membrana plasmatica2.
Abl prezinta o structura proteica complexa cu cel putin trei domenii functionale: un domeniu responsabil de fosforilare si celelalte doua, SH2 si SH3, care regleaza activitatea primului. Capacitatea Abl de transformare celulara este proportionala cu abilitatea sa de a fosforila reziduurile de tirozina. Se presupune ca Abl se leaga de proteina-tinta; domeniul SH2 creste activitatea kinazica promovand transformarea celulara, in timp ce domeniul SH3 reduce activitatea Abl.
In plus, se crede ca SH3 ar interveni si in reglarea activitatii GTP-azice, cunoscuta a fi implicata in calea de transductie a semnalului unei alte proto-oncogene, ras, a carei expresie anormala este asociata cu dezvoltarea cancerului. Sunt descrise si alte doua domenii functionale la nivelul proteinei Abl: un domeniu care se leaga de secvente nucleotidice specifice ale ADN-ului, ceea ce sugereaza posibilitatea ca Abl sa fie si un factor de transcriptie; in al doilea rand exista o regiune care faciliteaza legarea de F-actina in citoplasma.
Prin studii a fost demonstrat faptul ca pierderea regiunii N-terminale normale a Abl care mediaza autoreglarea, ca urmare a translocatiei Ph, conduce la cresterea activitatii de fosforilare a tirozinei precum si la o legare crescuta a F-actinei, transformand astfel Abl intr-o proteina oncogenica ce ar putea fi responsabila de proliferarea anormala a precursorilor mieloizi in LMC8;13.
Gena BCR are o lungime de 130 kb si contine 23 de exoni. Primul intron care separa exonii 1 si 2 a fost considerat initial ca avand o lungime de 68 kb, acum se stie insa ca include doi exoni aditionali (un exon alternativ e1’ si un exon alternativ e2’). Au fost identificati doi produsi de transcriptie de 4.5 si, respectiv, 7.0 kb in functie de modul de splicing alternativ; pot rezulta astfel 2 proteine de dimensiuni diferite, 130 si, respectiv, 160 kDa6. Proteina Bcr este exprimata in stadiile primare ale diferentierii mieloide, nivelul sau scazand in celulele mature; potrivit unor cercetatori ar putea constitui o serin/treonin kinaza mai degraba decat o tirozin kinaza ca proteina ABL8.
Regiunea 5’ netranslatata a genei are o importanta majora deoarece gena de fuziune rezultata in urma translocatiei Ph este reglata de promotorul BCR. Bogata in bazele nucleotidice G si C, aceasta regiune faciliteaza formarea de structuri loop si stem cu posibil rol in reglarea translationala, comuna in multe gene ‘housekeeping’ (gene exprimate constant in toate celulele)5;12.
Gena este exprimata ubiquitar, cu un nivel ridicat al ARN-ului mesager in tesutul hematopoetic si creier. Primul exon are o importanta majora, fiind inclus in toti produsii de fuziune BCR-ABL. La nivelul acestuia se exprima activitatea de serin/treonin kinaza a proteinei Bcr; astfel Bcr isi poate autofosforila reziduurile de serina si treonina. In afectiunile Ph pozitive, fosfotirozinele sunt prezente in ambele proteine Bcr si Bcr-Abl datorita activitatii de tirozin-kinaza a Bcr-Abl. Primul exon al componentei Bcr din Bcr-Abl este de asemenea fosforilat la reziduurile de serina si treonina.
In plus, acesta contine mai multe domenii care leaga regiunile SH2 ale altor proteine si detine in acest sens un rol important in asamblarea complexelor de transductie a semnalului. Un astfel de domeniu SH2 care se leaga de Bcr este cel al proteinei Abl, iar aceasta interactiune este mediata de catre serina si treonina fosforilata. Secventele Bcr implicate in aceasta interactiune sunt situate intre aminoacizii 192-242 si 298-413 si sunt esentiale in activarea oncogenica a Bcr-Abl. Un alt domeniu functional in exonul 1 al Bcr este cel de oligomerizare prin care s-a constatat ca Bcr si Bcr-Abl co-imunoprecipita.
Domeniul de oligomerizare afecteaza de asemenea localizarea proteinei Bcr-Abl4;13. Desi proteina Abl normala poate fi identificata atat in nucleu cat si in citoplasma, s-a constatat ca Bcr-Abl este localizata numai in citoplasma si este partial asociata cu citoscheletul. Deletia domeniului de oligomerizare al Bcr este insotita de reducerea legarii Bcr-Abl de F-actina, demonstrand ca acest domeniu intensifica capacitatea de legare a F-actinei de catre Bcr-Abl si este cel putin in parte responsabil de localizarea citoplasmatica a oncoproteinei15.
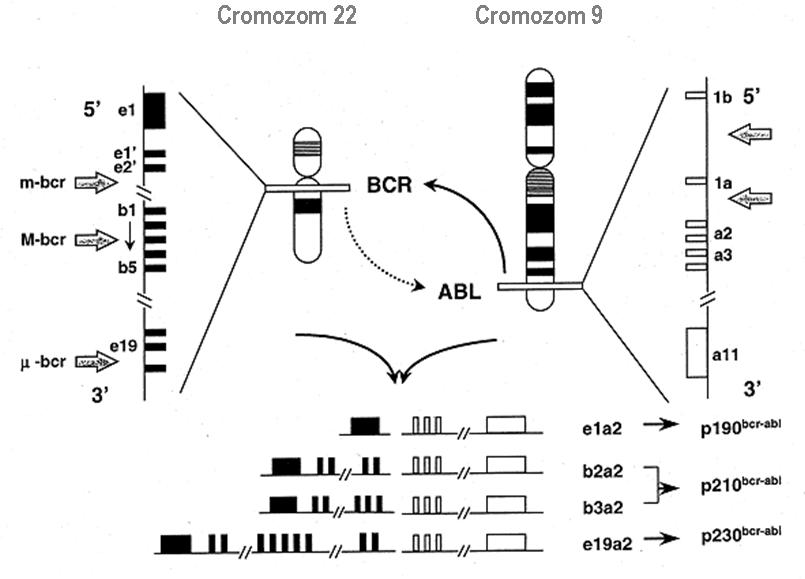
La nivelul ADN-ului, fuziunea BCR-ABL1 se caracterizeaza printr-o heterogenitate a punctelor de ruptura („breakpoints”) si o variatie a produsilor de transcriptie obtinuti printr-un splicing alternativ. Astfel, punctele de ruptura ale genei BCR de pe cromozomul 22 se gasesc in trei regiuni diferite: majora (M-bcr), minora (m-bcr) si μ-bcr; regiunea M-bcr este relativ scurta (5.8 kb), include 5 exoni denumiti b1-b5 (reprezinta de fapt exonii e12-e16 ai genei BCR), iar punctele de ruptura sunt localizate specific intre exonii b2 si b3 sau b3 si b4; m-bcr include exonii e1, e2, e1’ si e2’; μ-bcr include exonii e19 si e20. Pe de alta parte, punctele de ruptura al genei ABL sunt distribuite pe o lungime de 300 kb, intre capatul 5’ si exonul 2.
In functie de localizarea punctelor de ruptura segmente de dimensiuni diferite ale genei BCR fuzioneaza cu secvente 3’ ale genei ABL; aceste gene de fuziune vor genera 4 produsi de transcriptie – moleculele ARNm e1a2, b2a2, b3a2 si e19a2 – care in final vor conduce la sinteza a 3 proteine himerice cu greutati moleculare variabile (p190, p210 si respectiv p230) si, probabil, cu functii diferite (vezi figura 2).
Fig.2 Punctele de ruptura a genelor BCR si ABL (marcate prin sageti) si produsii de transcriptie rezultati
(Adaptare dupa Chasseriau et al, JMD, November 2004, vol.6, no.4)
Luand in considerare aceste date, se pot distinge cel putin 3 entitati clinico-patologice in LMC: p210 LMC, p190 LMC si p230 LMC. Marea majoritate a cazurilor de LMC este asociata cu produsii de transcriptie b2a2 si b3a2 care conduc la sinteza proteinei p210; desi in cele mai multe cazuri este prezent un singur tip de transcript, ocazional tumorile pot prezenta un splicing alternativ si produce simultan ambii produsi de transcriptie.
Proteina himerica p190 este detectata in ~50% din cazurile de LAL Ph pozitive, precum si in cazuri rare de LMC si LAM. Proteina himerica de greutate mai mare (p230) este prezenta in forme rare de LMC caracterizate printr-o componenta neutrofilica proeminenta; aceste cazuri nu trebuie incluse insa in entitatea „leucemie neutrofilica cronica”, ci mai degraba considerate ca fiind LMC, datorita prezentei genei de fuziune BCR-ABL.
Restul cazurilor de LAL Ph pozitive demonstreaza prezenta produsilor de transcriptie b3a2 sau b2a2. Merita sa fie mentionat faptul ca au fost identificate puncte de ruptura ale genei BCR in afara regiunilor mentionate in unele cazuri rare de LMC si LAL. In plus translocatia (9,22) a fost foarte rar detectata la pacienti cu alte neoplazii hematologice, cum ar fi mielomul multiplu si limfoamele cu celule B3;8;10.
Semnificatia clinica a diferitelor puncte de ruptura in LMC nu este clar definita. Cu toate acestea s-au putut efectua anumite corelatii. LMC survenita atat la copil cat si la adult este asociata aproape intotdeauna cu proteina himerica p210; in timp ce doua treimi din adulti prezinta transcriptul b3a2, la copii predomina b2a2. Mai mult, doua grupuri diferite de cercetatori au raportat asocierea b3a2 cu un numar mai mare de trombocite decat b2a2.
Aceasta constatare nu a fost confirmata insa de un al treilea grup din Marea Britanie8. In cazurile de LMC asociate cu transcriptul e1a2 componenta monocitara este mai proeminenta, iar numarul total de leucocite, bazofilia si splenomegalia sunt mai reduse decat in p210 LMC10.
Gene de fuziune BCR-ABL pot fi detectate si in leucocitele unor persoane sanatoase; in timp ce BCR-ABL se exprima relativ frecvent in celulele hematopoietice, doar foarte rar celulele dobandesc modificarile necesare pentru a produce leucemie3;10.
Activitatea tirozin kinazica este esentiala pentru procesele de semnalizare celulara si de crestere, iar intensificarea acesteia a fost asociata cu modificari oncogenice in cateva sisteme. Atat p210Bcr-Abl cat si p190Bcr-Abl prezinta activitate tirozin-kinazica constitutiva (constanta indiferent de cerintele fiziologice), cu niveluri mai mari inregistrate la proteina p190Bcr-Abl.
Majoritatea tirozinelor autofosforilate se gasesc la nivelul segmentului Bcr din Bcr-Abl, iar activitatea tirozin kinazica este atribuita domeniului kinazic localizat in segmentul Abl. Se crede ca gradul activitatii de transformare a Bcr-Abl se coreleaza cu gradul activitatii tirozin-kinazei si ca aceasta activitate a fost implicata in autonomia fata de factorii de crestere pe care Bcr-Abl o confera celulelor. De asemenea Bcr-Abl induce fosforilarea multor proteine implicate in transductia semnalului, adeziunea celulara, proliferare si apoptoza4.
Clasic, sunt descrise 2 sau 3 stadii clinice de evolutie a bolii, insa in practica medicala curenta a devenit din ce in ce mai frecvent ca pacientii sa fie diagnosticati in stadiul preclinic ca urmare a cresterii numarului controalelor periodice. Studiile au aratat ca exista o perioada de aproximativ 6 ani din momentul aparitiei translocatiei cromozomiale si dezvoltarea manifestarilor clinice.
Primul stadiu clinic este reprezentat de o faza cronica care in trecut avea o durata medie de 4 ani, urmata de o faza mai agresiva – faza accelerata/faza blastica cu un prognostic rezervat. Faza accelerata se caracterizeaza prin cresterea numarului de blasti in sangele periferic si/sau maduva osoasa, bazofilie, trombocitopenie persistenta, cresterea dimensiunilor splinei si a numarului de leucocite in conditiile tratamentului adecvat, evidentierea unei evolutii clonale la examenul citogenetic.
Faza blastica include urmatoarele elemente: ≥20% blasti in sangele periferic si/sau maduva osoasa, proliferarea extramedulara a blastilor sau prezenta unor aglomerari mari de blasti in maduva osoasa. Aceasta faza denota transformarea LMC in leucemie acuta, care poate fi de linie mieloida (~70%) sau limfoida (~30%).
Diagnosticul CML se bazeaza pe examenul frotiului de sange periferic, biopsia de maduva osoasa, examenul citogenetic pentru detectarea cromozomului Philadelphia si testele moleculare pentru evidentierea genei de fuziune BCR-ABL1 sau a produsilor de transcriptie asociati acesteia8.
Tratamentul definitiv al pacientilor cu LMC este transplantul de maduva osoasa alogenic sau transplantul de celule stem. Cu toate acestea, dezvoltarea inhibitorilor de tirozin kinaza cu introducerea pe scara larga a preparatului imatinib mesylate (Gleevec) a modificat semnificativ tratamentul de prima linie al LMC si a demonstrat succesul terapiei cu tinta moleculara3. Imatinibul, un inhibitor de tirozin kinaza specific pentru Abl, inhiba proliferarea liniilor celulare din LMC prin suprimarea activitatii kinazice a Bcr-Abl8.
Detectia prin RT-PCR a produsilor de transcriptie ARNm hibrizi BCR-ABL1 prezinta valoare in diagnosticul LMC si LAL Ph pozitive. Desi 98% dintre pacientii cu LMC sunt BCR-ABL pozitivi, intr-un numar redus de cazuri cu morfologie sugestiva pentru LMC marker-ul genetic lipseste. Aceste cazuri incadrate ca LMC atipica, leucemie mielo-monocitara cronica sau ca alte neoplazii mieloproliferative sau sindroame mielodisplazice beneficiaza de alte optiuni terapeutice decat cele de LMC Ph pozitive.
Pe de alta parte, sunt posibile manifestari atipice in LMC, iar in aceste cazuri demonstrarea anomaliei BCR-ABL este critica pentru diagnosticul corect si terapia adecvata. De asemenea testele moleculare au valoare diagnostica in situatiile in care examenul citogenetic este negativ pentru cromozomul Philadelphia (~5% din cazuri) ca urmare a unei anomalii BCR-ABL criptice sau submicroscopice. Detectia marker-ului la adultii si copiii cu LAL identifica acea categorie de pacienti care prezinta un risc crescut de esec terapeutic si care ar putea beneficia de programe de tratament intensiv3;8.
Un alt rol al testelor moleculare este monitorizarea raspunsului terapeutic la pacientii cu LMC. Astfel, dupa obtinerea remisiunii citogenetice sub Imatinib testele PCR cantitative reprezinta metoda de electie pentru detectarea bolii minime reziduale (MRD)8. De asemenea acestea isi gasesc aplicatia si la pacientii care au primit transplant medular pentru identificarea precoce a recurentelor bolii10.
Administrarea de Imatinib reprezinta tratamentul standard al pacientilor cu LMC, iar ELN (European LeukemiaNet) si NCCN (National Comprehensive Cancer Network) recomanda o doza zilnica de 400 mg ca optiune terapeutica de prima linie. Pacientii care obtin un raspuns citogenetic complet necesita monitorizarea raspunsului molecular la 3-6 luni pentru identificarea cazurilor de raspuns suboptimal si imbunatatirea tratamentului prin administrarea unor doze mai mari de Imatinib sau a unor terapii alternative.
De asemenea monitorizarea prin teste moleculare permite recunoasterea precoce a rezistentei primare sau dobandite la Imatinib. Inhibitorii de tirozin kinaza de generatia a doua, cum ar fi nilotinib si nisatinib, actioneaza mai eficient asupra tintei moleculare BCR-ABL fiind asociate cu o rata mai mare de raspuns citogenetic si molecular precoce; cuantificarea BCR-ABL ARNm va avea astfel o importanta si mai mare in viitorul apropiat.
Raspunsul molecular major (MMR) este definit prin scaderea nivelului BCR-ABL cu cel putin 3 log (de 1000 ori) fata de o valoare bazala standardizata (standardized baseline). Aceasta valoare a fost stabilita prin procesarea in 3 laboratoare centrale implicate in studiul IRIS (International Randomized Study of Interferon versus STI571) a unor esantioane efectuate din 30 probe provenite de la pacienti cu LMC inainte de initierea terapiei11.
Fiecare laborator a obtinut o valoare mediana pentru nivelul produsilor de transcriptie in cele 30 probe analizate care a constituit valoarea bazala standardizata specifica laboratorului. Reducerea cu > 3 log a nivelului BCR-ABL din aceasta valoare bazala standardizata specifica a definit raspunsul molecular major. Faptul ca s-a utilizat o valoare bazala absoluta (comuna tuturor participantilor) in loc de una relativa (individualizata) a asigurat faptul ca pacientii cu raspuns similar prezinta acelasi grad de boala reziduala.
Obtinerea unui MMR este asociata cu o probabilitate mai mare de raspuns pe termen lung si cu imbunatatirea perioadei de supravietuire fara progresia bolii (PFS = progression-free survival) la pacientii tratati cu Imatinib. Astfel, in studiul IRIS toti pacientii cu MMR la 12 luni de la inceputul tratamentului nu au prezentat nici un semn de faza accelerata sau criza blastica la evaluarea efectuata la 60 luni.
Este important de mentionat faptul ca in acest studiu a fost evitata folosirea termenului de „raspuns molecular complet“ fiind determinat numai numarul pacientilor la care s-a obtinut un rezultat nedetectabil al produsilor de transcriptie cu o sensibilitate analitica de 4.5 log sub valoarea bazala standardizata. Deoarece un rezultat nedetectabil reflecta limita de detectie a metodelor actuale, acesta nu trebuie considerat o dovada de eradicare a bolii16.
Avand in vedere importanta clinica a evaluarii prezentei bolii minime reziduale au fost initiate mai multe eforturi de a standardiza cuantificarea BCR-ABL. In acest sens, un grup de experti a propus la intalnirea internationala de consens de la Bethesda (2005) ca masuratorile BCR-ABL sa fie exprimate pe o scala internationala (IS) care foloseste doua valori standard: valoarea bazala standardizata (stabilita in studiul IRIS) considerata ca fiind 100% pe IS si valoarea MMR standardizata considerata ca fiind 0.1% pe IS (corespunde unei reduceri cu 3 log fata de valoarea bazala). O valoare de 1-2% IS corespunde in linii mari cu limita de detectie a metafazelor Ph1-pozitive prin examenul citogenetic standard. O valoare < 0.1% IS indica raspunsul molecular major (MMR)11.
Recomandari pentru efectuarea testului
- diagnosticul LMC si LAL Ph pozitive;
- monitorizarea raspunsului molecular la tratament si stabilirea prognosticului;
- detectarea bolii minime reziduale8.
Specimen recoltat – a) sange venos; b) aspirat medular9.
Recipient de recoltare – a) si b) vacutainer ce contine EDTA ca anticoagulant9.
Cantitate recoltata – a) 3-5 mL; b) 3 mL (minimum 1 mL)9.
Cauze de respingere a probei – folosirea heparinei ca anticoagulant9.
Stabilitate proba – maximum 3 zile la 2-8ºC9.
Metoda
Se folosesc doua tipuri de testari:
Detectia calitativa a produsilor de transcriptie ARNm BCR-ABL1 (in scop diagnostic)
Include urmatoarele etape:
- izolarea si purificarea ARN-ului;
- reactia de revers transcriptie (RT-PCR) pentru obtinerea ADN-ului complementar (ADNc) folosit ca matrita pentru amplificarea PCR;
- multiplex PCR (detectia uneia sau mai multor secvente ADN tinta in amestec prin folosirea unuia sau a mai multor seturi de primeri oligonucleotidici) + doua nested-PCR optimizate pentru detectarea diferitelor puncte de ruptura din genele BCR si ABL;
- fiecare rulare a probelor este insotita de o reactie de control cu ADNc de la o linie celulara cu translocatia t(9;22) si o reactie de control negativ cu ADNc al unei linii celulare care nu prezinta translocatia; pentru controlul calitatii ARN din proba si a eficientei reactiei de revers-transcriptie se utilizeaza in paralel o amplificare RT-PCR a unui fragment ADNc al genei ABL umane;
- detectia produsilor PCR prin electroforeza in gel de agaroza;
- limita de detectie a testului este de cel putin 1 celula LMC la 100 000 celule normale1;7;9.
Detectia cantitativa a produsilor de transcriptie ARNm BCR-ABL1 (in scop de monitorizare)
Va fi determinata expresia produsilor de fuziune (nr. copii BCR-ABL) raportata la expresia unei gene de control intern (ABL) prin Real-Time PCR cantitativ LightCycler cu o conversie finala in procente.
Raportarea si interpretarea rezultatelor
Detectia calitativa
Un rezultat negativ indica absenta unui transcript (ARNm) BCR-ABL.
Un rezultat pozitiv indica prezenta unui produs transcriptie BCR-ABL al carui tip va fi comunicat9.
Detectie cantitativa
Rezultatul va fi exprimat ca procent BCR-ABL din totalul ABL, astfel:
[Nr. copii BCR-ABL/ Nr. copii ABL] x 100
Nivelul produsilor de transcriptie BCR-ABL reflecta numarul celulelor leucemice reziduale.
O reducere progresiva a nivelului produsilor de transcriptie in cursul tratamentului cu Imatinib reprezinta criteriul actual pentru raspunsul molecular in leucemia mieloida cronica.
MMR va fi raportat in conformitate cu studiul IRIS.
O crestere de 5-10 ori a nivelului produsilor de transcriptie (0.5 sau 1 log) a fost propusa ca valoare prag pentru recaderea moleculara sau rezistenta la tratament9;16.
Bibliografie
1. B Gleissner, H Rieder, E Thiel, C Fonatsch, L A J Janssen, B Heinze, J W G Janssen, C Schoch, N Goekbuget, J Maurer, D Hoelzer and C R Bartram. Prospective BCR-ABL analysis by polymerase chain reaction (RT-PCR) in adult acute B-lineage lymphoblastic leukemia: reliability of RT-nested-PCR and comparison to cytogenetic data. In Leukemia, December 2001, Volume 15, Number 12, Pages 1834-1840.
2. Chissoe, S. L.; Bodenteich, A.; Wang, Y.-F.; Wang, Y.-P.; Burian, D.; Clifton, S. W.; Crabtree, J.; Freeman, A.; Iyer, K.; Jian, L.; Ma, Y.; McLaury, H.-J.; Pan, H.-Q.; Sarhan, O. H.; Toth, S.; Wang, Z.; Zhang, G.; Heisterkamp, N.; Groffen, J.; Roe, B. A. Sequence and analysis of the human ABL gene, the BCR gene, and regions involved in the Philadelphia chromosomal translocation. In Genomics 27: 67-82, 1995.
3. David S. Viswanatha, Richard S. Larson. Molecular Diagnosis of Hematopoietic Neoplasms. In Henry’s Clinical Diagnosis and Management by Laboratory Methods, Saunders-Elsevier, 21st Edition, 2007, 1296-1299.
4. Eunice Laurent, Moshe Talpaz, Hagop Kantarjian and Razelle Kurzrock. The BCR Gene and Philadelphia Chromosome-positive Leukemogenesis. In Cancer Research, 61, 2343-2355, March 15, 2001.
5. Hariharan I. K., Adams J. M. cDNA sequence for human BCR, the gene that translocates to the ABL oncogene in chronic myeloid leukaemia. In EMBO J., 6: 115-119, 1987.
6. Heisterkamp N., Knoppel E., Groffen J. The first BCR gene intron contains breakpoints in Philadelphia chromosome positive leukemia. In Nucleic Acids Res., 16: 10069-10081, 1988.
7. Janna Yu Sidorova, Liudmila B Saltykova, Anton A Lyschov, Andrey Yu Zaritskey,Kudrat M Abdulkadyrov, Michail N Blinov. A rapid RT-PCR based method for the detection of BCR-ABL translocation. In Clin Pathol: Mol Pathol 1997;50:266-268.
8. Kaaren K. Reichard, Richard S. Larson, Ian Rabinowitz. Chronic Myeloid Leukemia. In Wintrobe`s Clinical Hematology, Greer J, Foerster J, Lukens J, Rodgers G, Pareskevas F, Glader B, 12th ed, Lippincott Williams & Wilkins, 2009, 2006-2015.
9. Laborator Synevo. Referintele specifice tehnologiei de lucru utilizate, 2010. Ref Type: Catalog.
10. Marshall A. Lichtman, Jane L. Liesveld. Chronic Myelogenous Leukemia and Related Disorders. Williams Hematology, Lichtman M, Beutler E, Kipps T, Seligsohn U, Kaushansky K, Prchal J., 7th ed, McGraw-Hill Medical, 2006, 1237-1248.
11. M. C. Müller, N.C. P Cross, P. Erben, T. Schenk, B. Hanfstein, T. Ernst, R. Hehlmann, S. Branford, G. Saglio, A. Hochhaus. Harmonization of molecular monitoring of CML therapz in Europe. In Leukemia (2009) 23, 1957-1963.
12. Muller A. J., Witte O. N. The 5′ noncoding region of the human leukemia-associated oncogene BCR/ABL is a potent inhibitor of in vitro translation. In Mol. Cell. Biol., 9: 5234-5238, 1989.
13. Pluk, H.; Dorey, K.; Superti-Furga, G.: Autoinhibition of c-Abl. In Cell 108: 247-259, 2002.
14. Saglio G. Molecular Biology-measuring and reporting Bcr-Abl Transcripts. Workshop on RQ-PCR for monitoring CML patients treated with imatinib, Orbassano, 2006.
15. Wetzler M., Talpaz M., Van Etten R. A., Hirsh-Ginsberg C., Beran M., Kurzrock R. Subcellular localization of Bcr, Abl, and Bcr-Abl proteins in normal and leukemic cells and correlation of expression with myeloid differentiation. In J.Clin. Investig., 92: 1925-1939, 1993.
16. www.eutos.org. European Leukemia Network. Imatinib PCR testing for CML.
Produsul a fost adăugat în coș
În plus, ai la dispoziție 30 de zile pentru a veni la recoltare.