înapoi la lista
TIPAJ HLA-A (PCR)
Inchide
Informatii generale TIPAJ HLA-A (PCR)
Genetica complexului major de histocompatibilitate
Complexul major de histocompatibilitate (MHC) reprezinta un grup de gene situat pe bratul scurt al cromozomului 6, care produce markeri celulari de suprafata ce detin un rol important in procesul de recunoastere a self-ului: molecule de clasa I si de clasa II. La om moleculele MHC de clasa I includ antigenele leucocitare (HLA = human leukocyte antigen) HLA-A, HLA-B si HLA-C, in timp ce moleculele MHC de clasa II includ HLA-DR, HLA-DQ si HLA-DP. Aceste molecule constituie antigenele clasice implicate in transplant. La nivelul MHC mai sunt codificate si molecule de clasa III: componentele legate de MHC ale complementului (C2, C4 si Bf), 21-hidroxilaza (CYP21), proteina de soc termic (Hsp) 70 si factorul de necroza tumorala (TNF).
MHC se intinde pe o lungime de 3600 kb si include 224 de gene identificate, dintre care este prevazuta exprimarea unui numar de 128; aproximativ 40% dintre genele exprimate au o functie legata de sistemul imun. Genele HLA sunt localizate in 6 sub-regiuni care se succed in urmatoarea ordine: HLA-A, HLA-C, HLA-B, HLA-DR, HLA-DQ, HLA-DP, HLA-A fiind distal de centromer. Fiecare sub-regiune codifica cel putin o glicoproteina de suprafata (vezi figura 1).
Figura 1: Succesiunea genelor HLA I si II pe cromozomul 6
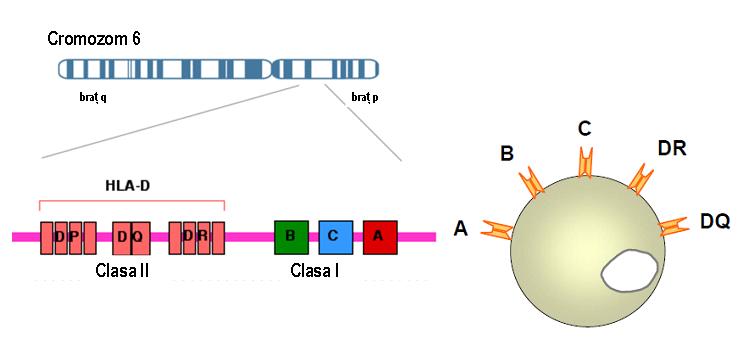
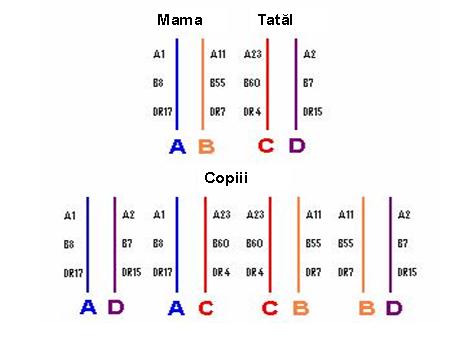
Cu o singura exceptie, genele HLA se caracterizeaza printr-un polimorfism inalt, mai exact fiecare gena prezinta alele multiple in populatie. Avand in vedere rolul important al sistemului HLA in raspunsul imun, se pare ca acest polimorfism este esential pentru supravietuirea speciilor si este mentinut in populatie prin selectie. Genele MHC sunt strans asociate intre ele (linkate), adica segrega in bloc in cursul meiozei. Complexul de gene linkate care sunt localizate pe una din perechile de cromozomi omologi si care segrega in bloc la urmasi este denumit haplotip. Fiecare individ mosteneste doua haplotipuri HLA – cate unul de la fiecare parinte – si are astfel doua alele pentru fiecare gena ce se exprima codominant (vezi figurile 2,3).
Figura 2: Alele-perechi mostenite de la mama si tata pentru fiecare locus HLA
Observatia ca alelele de pe diverse locusuri genetice apar in populatie in cadrul aceluiasi haplotip, cu o frecventa semnificativ mai mare decat cea prevazuta numai pe baza intamplarii, defineste dezechilibrul de linkaj – caracteristica sistemului HLA – care se intinde de la HLA-A pana la HLA-DQ inclusiv. Cel mai cunoscut dezechilibru de linkaj este haplotipul A1, Cw7, B8, DR17(3), DR52, DQ2 intalnit in populatia caucaziana cu o frecventa de 4 ori mai mare decat cea asteptata2. Figura 3: Modul de transmitere a haplotipurilor HLA la copiii unei familii

Rolul moleculelor HLA clasa I si II
Moleculele clasice apartinand clasei I MHC, denumite la om HLA-A, HLA-B si HLA-C, sunt heterodimeri alcatuiti dintr-un polipeptid transmembranar glicozilat asociat cu β2-microglobulina, ce se exprima pe toate celulele nucleate si detin roluri-cheie in raspunsul imun:
- Prezinta antigene peptidice derivate din proteine citoplasmatice (de exemplu, un virus sau o proteina anormala dintr-o celula maligna) receptorilor clonali specifici fiecarui peptid ai limfocitelor T CD8+. Aceste peptide rezulta in urma digerarii proteinelor la nivelul lizozomilor, avand in general o lungime de aproximativ 9 aminoacizi. Fiecare alotip MHC I isi selecteaza celulele T proprii; astfel exista 6 subseturi de celule T, fiecare fiind specializat pentru alotipurile HLA-A, HLA-B si HLA-C de provenienta materna sau paterna. In urma recunoasterii peptidelor celulele T CD8+ lizeaza celulele-tinta.
- Exprimarea lor pe suprafata celulelor exercita o functie protectoare in cadrul raspunsului imun innascut prin impiedicarea lizarii celulelor-tinta de catre celulele NK („natural killer”). In mod specific, NK pot recunoaste si liza celulele tinta care nu exprima pe suprafata lor moleculele HLA de clasa I, detinand astfel un rol important in supravegherea impotriva virusurilor si celulelor tumorale. Nivelul exprimarii moleculelor HLA de clasa I pe suprafata celulelor este maxim pe celulele limfoide; in cursul raspunsului imun exprimarea este crescuta sub actiunea citokinelor (interferon γ si TNF); pe de alta parte, tumorile si anumite virusuri (cum ar fi HIV) pot suprima exprimarea HLA I.
Alaturi de genele clasice, in regiunea MHC I mai sunt incluse gene cu un polimorfism redus desemnate ca HLA-E, HLA-F si HLA-G2;8.
Moleculele apartinand clasei II MHC, HLA-DR, HLA-DQ si HLA-DP, sunt heterodimeri alcatuiti din doua glicoproteine transmembranare asociate non-covalent – un lant α (33-35 kDa) si un lant β (26-28 kDa).
Fiecare din cele 3 sub-regiuni, DR, DQ si DP, include cel putin o gena A (care codifica lantul α) si o gena B (care codifica lantul β). Sub-regiunea DR codifica fie una, fie doua molecule DR, in functie de haplotip. Contine o singura gena DRA exprimata care este similara in mai multe haplotipuri. Gena DRB cea mai apropiata de centromer, DRB1, codifica un lant β extrem de polimorf care, asociat cu lantul α, formeaza molecula de clasa II ce predomina pe suprafata celulelor si care exprima specificitatile serologice DR1-DR18.
Cea de a doua gena DRB exprimata, care este prezenta doar in anumite haplotipuri, este localizata intre locusurile DRB1 si DRA si, in functie de alelele DRB1 exprimate, este denumita DRB3, DRB4 sau DRB5. Moleculele DR rezultate poarta specificitatile serologice DR52, DR53 si respectiv DR51. Sub-regiunea DQ include genele polimorfe DQA1 si DQB1 care codifica heterodimerul DQ ce poarta specificitatile serologice DQ1-9. Sub-regiunea DP contine 2 seturi de gene A si B: un set DPA1 si DPB1 cu polimorfism inalt, codifica produsul proteic DP; celalalt set este alcatuit din pseudogene2.
Spre deosebire de moleculele clasei I, cele din clasa II sunt exprimate numai pe suprafata celulelor prezentatoare de antigen (APC): monocite, macrofage, celule dendritice, limfocite B si, in plus, pe limfocitele T activate si celulele epiteliale timice8.
Moleculele clasei II prezinta limfocitelor T helper CD4+ fragmente antigenice peptidice procesate din antigene exogene, cum ar fi bacteriile. Receptorii pentru antigen ai limfocitelor T CD4+ interactioneaza cu complexul fragment antigenic-molecule HLA clasa II si declanseaza activarea celulelor. Ca si in cazul clasei I, recunoasterea fragmentului antigenic de catre celulele T efectoare este influentata de polimorfismul alelelor MHC.
Astfel, celula T efectoare este specifica pentru peptidul antigenic in conjunctie cu produsul alelic particular de clasa II care a prezentat initial antigenul (restrictie MCH). In urma recunoasterii antigenului fiecare celula T CD4+ activata stimuleaza diferentierea celulelor B in plasmocite producatoare de anticorpi si faciliteaza diferentierea altor limfocite T in celule citotoxice sau cu functii supresoare.
Figura 7: Prezentarea fragmentelor antigenice celulelor T de catre moleculele HLA I si II exprimate pe APC supresoare.
Figura 7: Prezentarea fragmentelor antigenice celulelor T de catre moleculele HLA I si II exprimate pe APC
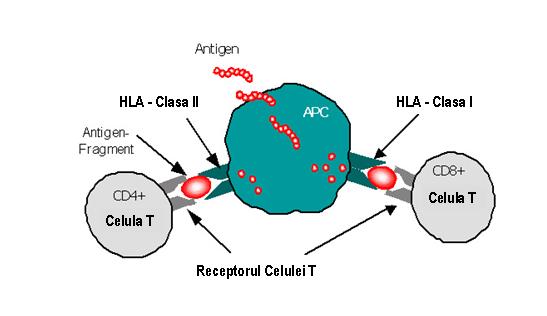
Atat moleculele de clasa I cat si cele de clasa II pot lega alternativ peptide proprii („self”) provenite din degradarea fiziologica a proteinelor celulare. In mod obisnuit acestea nu declanseaza activarea limfocitelor T deoarece sunt tolerate de sistemul imun individual; in unele situatii insa se pot declansa reactii imune care initiaza un proces de distrugere a self-ului ce conduce la autoimunitate2. Nomenclatura HLA Nomenclatura HLA este elaborata de catre un comitet OMS constituit in acest sens. Identificarea initiala a moleculelor HLA a implicat folosirea de aloseruri umane obtinute ca urmare a aloimunizarii produse in cursul sarcinii sau a efectuarii diverselor transplante. Astfel, nomenclatura rezultata reflecta istoria acestui efort de descoperire, aceasta fiind stabilita in cadrul unor workshop-uri internationale. Alotipurile au fost desemnate prin numere, cum ar fi HLA-A2, HLA-B27 si HLA-Cw6, iar numerele antigenelor HLA-A si HLA-B nu se suprapun (vezi figura 4). Litera „w” folosita initial pentru a desemna o anume categorie de workshop a fost pastrata pentru alotipurile HLA-C cu scopul de a le distinge de componentele complementului C2 si C4 care de asemenea sunt mapate in cadrul MHC8. Figura 4: Tipaj HLA serologic 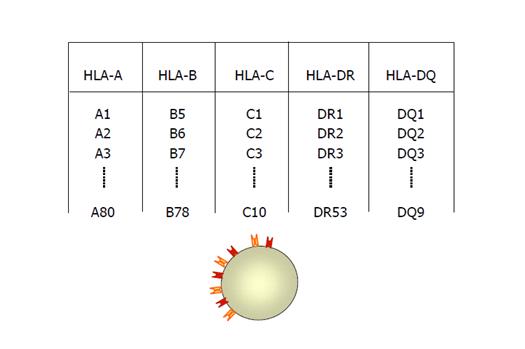
La testarea serologica unele aloseruri interactioneaza cu mai mult de un produs alelic HLA, fenomen denumit reactivitate incrucisata. In cazul produsilor alelici corespunzatori locusurilor HLA-A si HLA-B acest fenomen a fost mult studiat si utilizat pentru a clasifica moleculele in grupuri antigenice care reactioneaza incrucisat (CREGs). Moleculele de clasa I din cadrul unui CREG au in comun unul sau mai multi determinanti antigenici care nu se regasesc la nivelul moleculelor unui alt CREG. Un determinant antigenic (epitop) care este intalnit la membrii unui CREG este denumit specificitate publica (de exemplu: HLA-Bw4 si HLA-Bw6)2. Metodele serologice de tipaj HLA sunt inlocuite din ce in ce mai mult cu metode de biologie moleculara (electroforeza de acizi nucleici, tehnici de hibridizare, precum si de secventiere), care au condus la modificarea nomenclaturii prin luarea in considerare a alelelor ADN. Astfel, o singura specificitate definita serologic poate fi exprimata de doua sau mai multe (>25) alele diferite (vezi figura 5).
Figura 5: Tipaj HLA – de la serologie la biologie moleculara
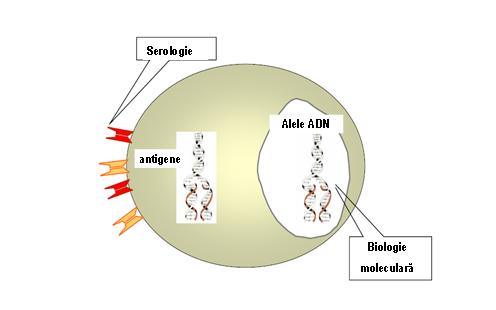
In cadrul nomenclaturii bazate pe ADN fiecare alela HLA este desemnata prin numele locusului genei urmat de un asterisc * si un numar de 4-7 cifre care indica alela (de exemplu, A*0201 este o alela a genei HLA-A si B*2701 este o alela a genei HLA-B). Primele 2 cifre in denumirea fiecarei alele se bazeaza frecvent pe tipul serologic al moleculei rezultate si/sau similaritatea secventelor nucleotidice la alte alele din cadrul grupului (vezi figura 6). Figura 6: Tipaj HLA prin biologie moleculara


Unele alele difera prin secventa ADN a exonilor, dar nu si prin secventa de aminoacizi rezultata (datorita unor substitutii silentioase sau sinonime); aceste alele sunt identificate prin adaugarea unei a 5-a cifre la cele 4 cifre comune (de exemplu, B*27051 si B*27052). In plus, cifrele 5-7 adaugate in denumire indica faptul ca 2 alele difera numai prin secvente la nivelul intronilor (de exemplu, DRB4* 0103101 si DRB4* 0103102). In exemplul dat DRB4* 0103102 prezinta un situs de splicing al ARN-ului modificat care conduce la pierderea expresiei alelei; alelele care nu se exprima ca proteine pot avea un N adaugat la numele lor (de exemplu, A*215N si DRB4* 0103102N)2;3. Spre deosebire de tehnicile serologice, tipajul HLA bazat pe ADN prezinta net avantaje:
- este specific (fiecare reactiv pentru tipaj este clar definit si se bazeaza pe o secventa nucleotidica specifica);
- este flexibil (pe masura ce sunt descoperite alele noi pot fi preparati reactivi noi);
- este mai robust decat alte tehnici (nu necesita limfocite viabile);
- poate fi aplicat pe scara larga;
- poate face o discriminare intre alelele ADN care specifica proteine HLA ce nu pot fi diferentiate serologic; de exemplu, o persoana care prezinta alela DRB1* 0401 are acelasi tip serologic DR4 ca si un individ care este purtator al alelei DRB4* 04122.
Prin tehnicile moleculare sunt identificate la ora actuala 4447 alele HLA clasa I si II: 965 HLA-A, 1543 HLA-B, 626 HLA-C, 9 HLA-E, 21 HLA-F, 46 HLA-G, 855 HLA-DRB, 35 HLA-DQA1, 107 DQB1, 28 HLA-DPA1, 138 HLA-DPB3. Noille alele descoperite sunt prezentate in rapoarte periodice ale Comitetului OMS, iar secventele nucleotidice ale tuturor alelelor sunt depozitate intr-o baza de date computerizata (GenBank, EMBL, IMGT/HLA databases)2. Din aprilie 2010 intra in vigoare o modificare in nomenclatura HLA: ca urmare a numarului de alele HLA in crestere s-a decis sa se introduca „:” in denumirea alelor pentru a delimita campuri separate. Astfel:
A*01010101 → A*01:01:01:01
A*260101 →A*26:01:01
A*3301 → A*33:01
B*0808N → B*08:08N
DRB1*01010101 → DRB1*01:01:01:01
Sunt de asemenea preconizate si alte modificari, printre care si aceea ca litera „w” va fi scoasa din denumirea alelelor HLA-C, dar va fi mentinuta in cea a antigenelor HLA-C4.
Boli asociate cu sistemul HLA
Exista un numar semnificativ de afectiuni care prezinta o asociere cu genele MHC; in cele mai multe cazuri este vorba de boli autoimune care nu au un mod de transmitere mendelian clasic. De exemplu, spondilita ankilopoietica se asociaza in 95% din cazuri cu HLA-B27.
O problema majora in legatura cu asocierile de gene MHC este penetranta lor incompleta, ceea ce face ca studiile formale de segregare si linkaj sa fie foarte dificil, daca nu imposibil, de efectuat. O alta necunoscuta este numarul alelelor predispozante pentru o anumita boala intr-o populatie specifica; se pare ca multe boli asociate cu sistemul HLA sunt poligenice.
Asocierea genetica este reflectata in termenul de „risc relativ”, care reprezinta raportul statistic intre riscul individual de boala la purtatorii unui marker genetic si riscul la indivizii aceleiasi populatii care nu au acest marker. Un risc relativ mai mare decat 1 indica o asociere pozitiva intre HLA si boala respectiva6.
Printre afectiunile asociate cu HLA care au fost studiate extensiv se numara diabetul zaharat tip 1. Cea mai puternica asociere este cea cu alelele clasei II HLA DR si DQ. Genotipul heterozigot DR3/DR4 confera cel mai mare risc de diabet, urmat de genotipurile homozigote DR4 si DR3.
La caucazieni heterodimerii HLA-DQ (lanturile alfa DQA1 si lanturile beta DQB1) codificati de alelele DQA1*0301, DQB1*0302, DQA1*0501, DQB1*0201 prezinta cea mai mare asociere cu diabetul de tip 1. S-a observat ca DQB1*0302 difera de DQB1*0301 in pozitia 57, unde lipseste un rest de acid aspartic. Aceeasi lipsa a acidului aspartic in pozitia 57 apare si la alela DQB1*0201 si s-a sugerat ca restul de aminoacid ar fi implicat in mecanismul molecular al susceptibilitatii fata de diabet3.
Alte haplotipuri ale genelor HLA clasa II au rol protector fata de dezvoltarea diabetului: DQA1*0102/DQB1*0602/DRB1*1501. In special alela DQB1*0602 confera protectie, chiar si la rudele de gradul I ale pacientilor diabetici care prezinta anticorpi anti-celule insulare pancreatice9.
Riscul absolut pentru diabet zaharat tip 1 in functie de genotipul HLA este prezentat in tabelul de mai jos1:
| DR3/DR3 | 1/125 |
| DR3/DRX | 1/500 |
| DR4/DR4 | 1/147 |
| DR4/DRX | 1/476 |
| DR3/DR4 | 1/42 |
| DRX/DRX | 1/5565 |
Testarea HLA este utila pentru evaluarea riscului de a dezvolta diabet zaharat la persoane cu istoric familial pozitiv.
Pacientii purtatori ai markerilor HLA DQB1*0201 si/sau *0302 sau ai alelelor predispozante DR4 (DRB1*0405, DRB1*0402) ar trebui urmariti biochimic si imunologic5.
Ca urmare a introducerii tehnicilor bazate pe ADN in tipajul HLA au fost depistate cateva asocieri clare intre progresia infectiei HIV-1 catre sindromul imunodeficientei dobandite (SIDA) si unele antigene HLA clasa II. Astfel, in prezenta HLA-B27 se constata o absenta a progresiei catre SIDA pe termen lung, in timp ce asocierea cu HLA-B35 denota o progresie rapida. Diferenta intre cele 2 alele consta in numarul diferit de peptide virale care se pot lega de cele 2 molecule HLA-B exprimate; molecula HLA-B27 leaga 15 tipuri diferite de peptide provenite din anvelopa virusului, pe cand nici un peptid viral nu contine secvente de aminoacizi care sa se lege preferential de HLA-B35. Prin urmare, incarcarea moleculelor HLA cu peptide in vederea prezentarii antigenului constituie o etapa critica. Infectia HIV este controlata de catre celulele T CD8+ o perioada de timp variabila; marimea acestei perioade depinde de numarul peptidelor virale recunoscute8.
In ceea ce priveste asocierea HLA cu borrelioza, conform datelor din literatura sunt raportate:
- HLA asociate cu boala Lyme refractara la tratament, adica cu autoimunitate indusa de agentul patogen:
DR1 (HLA-DRB1*0101)
DR2 (HLA-DRB1*1501)
DR4 (HLA-DRB1*0401,*0402,*0403,*0404,*0405,*0407)
- HLA asociate cu productie scazuta de anticorpi specifici de Borrelia, in prezenta infectiei certe:
alele DR1 (HLA-DRB1*0101,*0102,*0103,*0104,*0105).
In concluzie, determinarea antigenelor HLA poate fi folosita astazi ca marker pentru diagnosticul diferenţial al multor afecţiuni. In tabelul de mai jos sunt prezentate exemple de afecţiuni asociate cu anumite constelaţii HLA, impreuna cu riscul relativ de boala7.
| Boala | Antigen HLA | Risc relativ |
| Alergia la latex | DR4, DQB1*0302 (DQ3) | 2.4 |
| Alergia la insulina | B7, B21 | 5.4 |
| Alopecia areata | B12 | 5.4 |
| Alveolita alergica exogen-allergischexogena | DR6 | 16.5 |
| Anemia aplastica | DR2 | 2.2 |
| Anemia pernicioasa | DR5 | 5.4 |
| Artrita cronica juvenila | DR8 DR5 | 8.0 3.3 |
| Artrita psoriazica | B27 B38 | 10.7 9.1 |
| Artrita reactiva – infectii cu: Shigella, Yersinia, Salmonella | B27 | 40.0 |
| Artrita Lyme (infectie cu Borrelia) | DR2/DR4 | 22.0 |
| Astm indus de aspirina | DQ2 | 4.1 |
| Boala Addison (idiopatica) | DR3 | 6.3 |
| Boala Alzheimer | B7/Cw3 | 28.0 |
| Boala Basedow | DR3 | 4 |
| Boala Behcet | B51 | 6.3 |
| Boala M. Behcet (mucokutane Form)Behcet (forma muco-cutanata) | B12 | 5.6 |
| Boala Bechterew | B27 | 87.4 |
| Boala celiaca | DQ2/DQ7/DQ8 | 52.0 |
| M. CrohnBoala Crohn | DR1, DR4, DR7 | 2.0-7.0 |
| Boala mixta de tesut conjunctiv | DR4 | 3 |
| Ciroza biliara primara (PBC) | DR3, DRB1*0803 (DR8) | 3-6.8 |
| Corioretinopatie “birdshot” | A29 | 224 |
| Cholangitis, primär sklerosierend (PSC)Colangita sclerozanta primitiva (CSP) | DR3, DR8 | 4.5-15 |
| Dermatita herpetiforma | DR3/DR7 | 17.3 |
| Dermatomiozita | DR3/DR52 | 5 |
| Diabet zaharat tip I | DQ2/DQ3 subtipuri DRB1*0405 (DR4)DQ*0602 (DQ6)DQB1*0602 (DQ6) | > 50 Protectiv |
| Deficit selectiv de IgA, | DR3 | 17.0 |
| Glomerulonefrita membranoasa idiopatica | DR3 | 12.0 |
| Granulomatoza Wegener | DR9 | 6.7 |
| Hemocromatoza idiopatica | A3/B7/B14 | 90.0 |
| Hepatita cronica autoimuna | DR3, DR4, DR8, DR52 | 4.7 |
| Hepatita B, evolutie cronica | B35 | 158.0 |
| Ag Hbs – purtatori sanatosi | B41 | 11.2 |
| Nebennieren-Rinden-Hyperplasie, kongenitalHiperplazia CSR congenitala | B47 DR3 | 15.4 6.3 |
| HIV – hipersensibilitate la Abacavir | B * 5701 | 117 |
| HIV – evolutie lenta | B27 B57 | Protector Protector |
| Lichen plan | DR1 | 11.8 |
| Litiaza biliara | A19 | 131 |
| Lupus eritematos sistemic (LES) | DR2/DR3 | 5.8 |
| Lupus iatrogen | DR4 | 5.6 |
| Miastenia gravis | DR3 DR5 | 3.9 3.6 |
| Narcolepsie | DQB1*0602, DRB1*1501 | 130.0 |
| Lupus-NephritisNefrita lupica | DR2/DQ1 | 14.0 |
| Nefropatia IgA | DR4 | 5.5 |
| Nefropatia membranoasa idiopatica | DR3 | 3-12 |
| Panencefalita sclerotica subacuta (SSPE) | A29 | 3.6 |
| Pemfigus vulgar | DR3, DR4 | 14.4 |
| Pemfigus foliaceu | DR1 | 7.3 |
| Poliartrita reumatoida | DR1/DR4 subtypes | 10.2 |
| Polimialgia reumatica | DR3 DR4 | 2.3 5.7 |
| Polimiozita | DR3/DR52 | |
| Psoriazis vulgar | B17, B37 Cw6 | 6.7 33.0 |
| Purpura trombocitopenica autoimuna | DR2 | 9.2 |
| Sindrom Reiter | B27 | 37.0 |
| Retinopatia diabetica | B8 | 4.0 |
| Sarcoidoza | B7/B8 DR2, DR3, DR5 | 8.5 |
| Sarcom Kaposi | DR5 | 5.3 |
| Schizofrenie | A9/B27 A2/A11 | 11.9 9.8 |
| Sclerodermie (PSS) | DR3 DR5 | 16.7 10,0 10.0 |
| Scleroza multipla | DR2/DQ6 subtypes | 4.1 |
| Sindrom androgenital – forma cu debut tardiv | B14 | 48.5 |
| Sindrom androgenital – forma cu pierdere de sare | B47 | 51.0 |
| Sindrom antifosfolipidic | DR4, DR7 | 5.1 |
| Sindrom astmatiform indus de analgezice | A1/B8/DR3 | 28.9 |
| Sindrom CREST | DR5 | 8.1 |
| Sindrome Goodpasture | DR2 | 15.9 |
| Sjögren-SyndromSindrom Sjögren | DR2/DR3 | 9.7 |
| Stomatita, aftoza recurenta | B7 | 6.0 |
| Tiroidita Hashimoto | DR3, DR5 | 3.2 |
| Tiroidita de Quervain (subacuta) | B35 | 13.7 |
| Tiroidita postpartum | DR4/DR5 | 8.0 |
| Trombocitopenia neonatala aloimuna | DR3 | 9.2 |
| Uveita anterioara acuta | B27 | 10-50 |
Pregatire pacient
nu este necesara o pregatire speciala; pe formularul de trimitere se va preciza afectiunea suspectata pentru care se solicita testarea HLA7.
Specimen recoltat – sange venos7.
Recipient de recoltare – vacutainer ce contine EDTA ca anticoagulant7.
Cantitate recoltata – cat permite vacuumul7.
Cauze de respingere a probei – folosirea heparinei ca anticoagulant; probe coagulate sau hemolizate; probe refrigerate sau congelate7.
Stabilitate proba – 7 zile la 2-8ºC7.
Metoda de lucru - reactie de polimerizare in lant (PCR) cu detectie prin electroforeza in gel de agaroza7.
Interpretarea rezultatelor
Rezultatul pozitiv la un antigen HLA asociat cu o anumita afectiune arata o predispozitie genetica. Rezulta un risc relativ al purtatorului de a se imbolnavi, comparativ cu populatia „normala” (vezi tabelul). In cazul suspiciunii unei anumite afectiuni, evidentierea antigenului HLA reprezinta un criteriu de diagnostic7.
Bibliografie
1. E. Albert Reece, Donal R.Coustan, Steven G.Gabe. Diabetes in Women, ed.3, 2004, 61-62.
2. H. Davis, Richard A. McPherson. Human Leukocyte Antigen: The Major Histocompatibility Complex of Man. In Henry’s Clinical Diagnosis and Management by Laboratory Methods, Saunders-Elsevier, 21st Edition, 8, 876-886.
3. HLA Nomenclature. IMGT/HLA database. www.ebi.ac.uk/imgt/hla/nomenclature/index.html. Ref Type: Internet Communication.
4. HLA Nomenclature Changes Effective April 1, 2010. www.ashi-hla.org. Ref Type: Internet Communication.
5. Jacob Sten Petersen. Antigen-based Prediction and Prevention of Type 1 Diabetes. In Danish Medical Bulletin nr.4/2006 418-437.
6. Julio C. Delgado, Edmond J. Yunis. The Major Histocompatibility Complex and Disease. In Henry’s Clinical Diagnosis and Management by Laboratory Methods, Saunders-Elsevier, 21st Edition, 8, 894-902.
7. Laborator Synevo. Referintele specifice tehnologiei de lucru utilizate. Ref Type: Catalog.
8. Robert J. Winchester. The Major Histocompatibility Complex. In Clinical Immunology. Principles and Practice, Mosby, Elsevier, Third Edition, 2008, 79-89.9. V. Radha, K.S.Vimaleswaran, R.Deepa, V. Mohan. The genetics of diabetes mellitus. In Indian J Med Res 117/2003 225-238.
Vezi tot conținutul
Vezi mai puțin